Information for the Public
Nobel Prize in Chemistry Winners
8 October 2003
Aquaporin Water Channels
 |
 |
| Peter Agre | Roderick MacKinnon |
All living matter is made up of cells. A single human being has as many as the stars in a galaxy, about one hundred thousand million. The various cells – e.g. muscle cells, kidney cells and nerve cells – act together in an intricate system in each one of us. Through pioneering discoveries concerning the water and ion channels of cells, this year’s Nobel Laureates Peter Agre and Roderick MacKinnon, have contributed to fundamental chemical knowledge on how cells function. They have opened our eyes to a fantastic family of molecular machines: channels, gates and valves all of which are needed for the cell to function.
Molecular channels through the cell wall
To maintain even pressure in the cells it is important that water can pass through the cell wall. This has been known for a long time. The appearance and function of these pores, remained for a long time as one of the classical unsolved problems of biochemistry. It was not until around 1990 that Peter Agre discovered the first water channel. Like so much else in the living cell, it was all about a protein.
Water molecules are not the only entities that pass into and out of the cell. For thousands of millions of cells to be able to function as something other than one large lump, coordination is required. Thus communication between the cells is necessary. The signals sent in and between cells consist of ions or small molecules. These start cascades of chemical reactions that cause our muscles to tense, our eyes to water – indeed, that control all our bodily functions. The signals in our brains also involve such chemical reactions. When we stub a toe this starts a signal moving up towards the brain. Along a chain of nerve cells, through interaction between chemical signals and ion currents, information is conveyed from cell to cell like a baton in a relay race.
It was in 1998 that Roderick MacKinnon succeeded for the first time in showing what ion channels look like at atomic level – an achievement which, together with Agre’s discovery of water channels, opened up entirely new research areas in biochemistry and biology.
The medical consequences of Agre’s and MacKinnon’s discoveries are also important. A number of diseases can be attributed to poor functioning in the water and ion channels of the human body. With the help of fundamental knowledge of what they look like and how they work, there are now new possibilities for developing new and more effective pharmaceuticals.
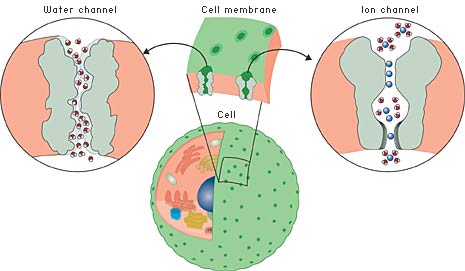 |
| Fig 1. The dividing wall between the cell and the outside world – including other cells – is far from being an impervious shell. On the contrary, it is perforated by various channels. Many of these are specially adapted to one specific ion or molecule and do not permit any other type to pass. Here to the left we see a water channel and to the right an ion channel. |
Water channels
The hunt for the water channels
As early as the middle of the nineteenth century it was understood that there must be openings in the cell membrane to permit a flow of water and salts. In the middle of the 1950s it was discovered that water can be rapidly transported into and out of cells through pores that admit water molecules only. During the next 30 years this was studied in detail and the conclusion was that there must be some type of selective filter that prevents ions from passing through the membrane while water molecules, which are uncharged, flow freely. Thousands of millions of water molecules per second pass through one single channel!
Although this was known, it was not until 1992 that anybody was able to identify what this molecular machinery really looked like; that is, to identify what protein or proteins formed the actual channel. In the mid-1980s Peter Agre studied various membrane proteins from the red blood cells. He also found one of these in the kidney. Having determined both its peptide sequence and the corresponding DNA sequence, he realized that this must be the protein that so many had sought before him: the cellular water channel.
Agre tested his hypothesis in a simple experiment (fig. 2) where he compared cells which contained the protein in question with cells which did not have it. When the cells were placed in a water solution, those that had the protein in their membranes absorbed water by osmosis and swelled up while those that lacked the protein were not affected at all. Agre also ran trials with artificial cells, termed liposomes, which are a type of soap bubble surrounded on the outside and the inside by water. He found that the liposomes became permeable to water if the protein was planted in their membranes.
| What is osmosis? The liquid pressure in plant and animal cells is maintained through osmosis. In osmosis, small molecules (such as water) pass through a semi-permeable membrane. If the membrane does not admit macromolecules or salts that are in higher concentrations on one side of the membrane, the small molecules (water) will cross to this side, attempting to ”dilute” the substance that cannot pass through the membrane. The osmotic pressure thus arising is the reason why cells are often swollen and stiff, in a flower stalk, for example. |
 |
| Fig 2. Peter Agre’s experiment with cells containing or lacking aquaporin. The aquaporin is necessary for making the cell absorb water and swell. |
Peter Agre also knew that mercury ions prevent cells from taking up and releasing water, and he showed that water transport through his new protein was prevented in the same way by mercury. This made him even more sure of that he had discovered what was actually the water channel . Agre named the protein aquaporin, ”water pore”.
How does the water channel work? A question of form and function
In 2000, together with other research teams, Agre reported the first high-resolution images of the three-dimensional structure of the aquaporin. With these data, it was possible to map in detail how a water channel functions. How is it that it only admits water molecules and not other molecules or ions? The membrane is, for instance, not allowed to leak protons. This is crucial because the difference in proton concentration between the inside and the outside of the cell is the basis of the cellular energy-storage system.
Selectivity is a central property of the channel. Water molecules worm their way through the narrow channel by orienting themselves in the local electrical field formed by the atoms of the channel wall. Protons (or rather oxonium ions, H3O+) are stopped on the way and rejected because of their positive charges.
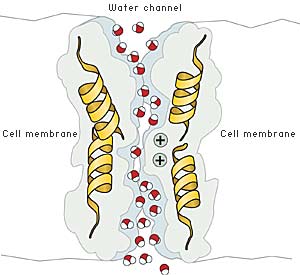 |
| Animations » |
| Fig 3. Passage of water molecules through the aquaporin AQP1. Because of the positive charge at the center of the channel, positively charged ions such as H3O+, are deflected. This prevents proton leakage through the channel. |
The medical significance of the water channels
During the past ten years, water channels have developed into a highly topical research field. The aquaporins have proved to be a large protein family. They exist in bacteria, plants and animals. In the human body alone, at least eleven different variants have been found.
The function of these proteins has now been mapped in bacteria and in plants and animals, with focus on their physiological role. In humans, the water channels play an important role in, among other organs, the kidneys.
The kidney is an ingenious apparatus for removing substances the body wishes to dispose of. In its windings (termed glomeruli), which function as a sieve, water, ions and other small molecules leave the blood as ‘primary’ urine. Over 24 hours, about 170 liters of primary urine is produced. Most of this is reabsorbed with a series of cunning mechanisms so that finally about one liter of urine a day leaves the body.
From the glomeruli, primary urine is passed on through a winding tube where about 70% of the water is reabsorbed to the blood by the aquaporin AQP1. At the end of the tube, another 10% of water is reabsorbed with a similar aquaporin, AQP2. Apart from this, sodium, potassium and chloride ions are also reabsorbed into the blood. Antidiuretic hormone (vasopressin) stimulates the transport of AQP2 to cell membranes in the tube walls and hence increases the water resorption from the urine. People with a deficiency of this hormone might be affected by the disease diabetes insipidus with a daily urine output of 10-15 liters.
Ion channels
The cells signal with salt!
The first physical chemist, the German Wilhelm Ostwald (Nobel Prize in Chemistry 1909) proposed in 1890 that the electrical signals measured in living tissue could be caused by ions moving in and out through cell membranes. This electro-chemical idea rapidly achieved acceptance. The notion of the existence of some type of narrow ion channel arose in the 1920s. The two British scientists Alan Hodgkin and Andrew Huxley made a major breakthrough at the beginning of the 1950s and for this were awarded the Nobel Prize in Physiology or Medicine in 1963. They showed how ion transport through nerve cell membranes produces a signal that is conveyed from nerve cell to nerve cell like a relay race baton. It is primarily sodium and potassium ions, Na+ and K+, that are active in these reactions.
Thus as much as fifty years ago there was well-developed knowledge of the central functions of the ion channels. They had to be able to admit one ion type selectively, but not another. Likewise, it had to be possible for the channels to open and shut and sometimes to conduct ions in one direction only. But how this molecular machinery really worked was long to remain a mystery.
Ion-selective channel
During the 1970s it was shown that the ion channels were able to admit only certain ions because they were equipped with some kind of ”ion filter”. Of particular interest was the finding of channels that admit potassium ions but not sodium ions – even though the sodium ion is smaller than the potassium ion. It was suspected that the oxygen atoms in the protein played an important role as “substitutes” for the water molecules with which the potassium ion surrounds itself in the water solution and from which it must free itself during entry to the channel.
But further progress with this hypothesis was difficult – what was now needed was simply high-resolution pictures of the type only X-ray crystallography can provide. The problem was that it is extremely difficult to determine the structure of membrane proteins with this method, and the ion channels were no exception. Membrane proteins from plants and animals are more complicated and difficult to work with than those from bacteria. Using bacterial channel proteins that resemble human ion channels as closely as possible might perhaps offer a way forwards.
Many researchers tried in vain. The breakthrough came from an unexpected direction. Roderick MacKinnon, after studying biochemistry, turned to medicine and qualified as medical doctor. After working as a physician for some years, he grew so interested in ion channels that he started to do research in the field: “My scientific career in effect began at the age of 30”, he has admitted. But his career took off quickly. Realizing that better and higher-resolution structures were needed for understanding how ion channels function, he decided to learn the fundamentals of X-ray crystallography. It was then only a few years before he astonished the whole research community by presenting a structure of an ion channel. This was in April 1998.
First ion channel mapped – atom by atom
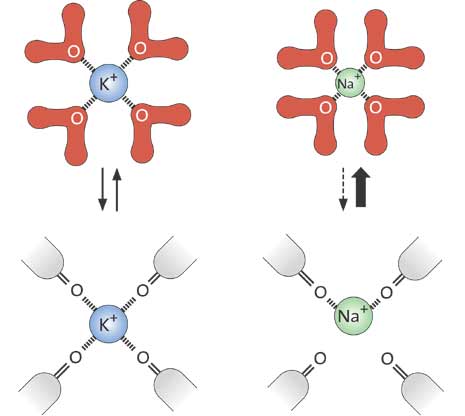
In 1998, then, MacKinnon determined the first high-resolution structure of an ion channel, called KcsA, from the bacterium Streptomyces lividans. MacKinnon revealed for the first time how an ion channel functions at atomic level. The ion filter, which admits potassium ions and stops sodium ions, could now be studied in detail. Not only was it possible to unravel how the ions passed through the channel, they could also be seen in the crystal structure – surrounded by water molecules just before they enter the ion filter; right in the filter, and when they meet the water on the other side of the filter (fig. 4). MacKinnon could explain why potassium ions but not sodium ions are admitted through the filter: namely, because the distance between the potassium ion and the oxygen atoms in the filter is the same as that between the potassium ion and the oxygen atoms in the water molecules surrounding the potassium ion when it is outside the filter. Thus it can slide through the filter unopposed. However, the sodium ion, which is smaller than the potassium ion, can not pass through the channel. This is because it does not fit between the oxygen atoms in the filter and therefore remains in the water solution. The ability of the channel to strip the potassium ion of its water and allow it to pass at no cost in energy is a kind of selective catalyzed ion transport.
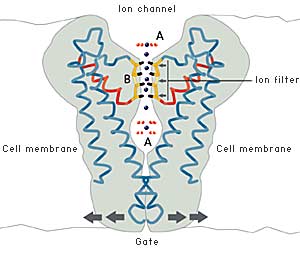
The cell must also be able to control the opening and closing of ion channels. MacKinnon has shown that this is achieved by a gate at the bottom of the channel which opened and closed a molecular “sensor”. This sensor is situated close to the gate. Certain sensors react to certain signals, e.g. an increase in the concentration of calcium ions, an electric voltage over the cell membrane or binding of a signal molecule of some kind. By connecting different sensors to ion channels, nature has created channels that respond to a large number of different signals.
| Fig 4. The ion channel permits passage of potassium ions but not sodium ions. The oxygen atoms of the ion filter form an environment very similar to the water environment outside the filter. The cell may also control opening and closing of the channel. |
 |
| High resolution image (jpeg 137 kB) » |
| OUTSIDE THE ION FILTER (A) Outside the cell membrane the ions are bound to water molecules with certain distances to the oxygen atoms of the water. |
 |
|
| High resolution image (jpeg 121 kB) » |
| INSIDE THE ION FILTER (B) For the potassium ions the distance to the oxygen atoms in the ion filter is the same as in water. |
The sodium ions, which are smaller, do not fit in between the oxygen atoms in the filter. This prevents them from entering the channel. |
Understanding diseases
Membrane channels are a precondition for all living matter. For this reason, increased understanding of their function constitutes an important basis for understanding many disease states. Dehydration of various types, and sensitivity to heat, are connected with the efficancy of the aquaporins. The European heat waves of recent years, for example, resulted in many deaths where the cause has sometimes been connected to problems in maintaining the body-fluid balance. In these processes the aquaporins are of crucial importance.
Disturbances in ion channel function can lead to serious diseases of the nervous system as well as the muscles, e.g. the heart. This makes the ion channels important drug targets for the pharmaceutical industry.
Illustrations: Typoform
Source: http://nobelprize.org/nobel_prizes/chemistry/laureates/2003/public.html
| The Laureates | |
| Peter Agre Departement of Biological Chemistry 420 Physiology Building Johns Hopkins University School of Medicine 725 North Wolfe Street Baltimore Maryland 21205 USA |
US citizen. Born 1949 (54 years) in Northfield, Minnesota, USA. B.A. at Augsburg College, Minneapolis, chemistry major, 1970. M.D. at Johns Hopkins University School of Medicine in Baltimore, 1974. Since 1993, Professor of Biological Chemistry and Professor of Medicine at Johns Hopkins School of Medicine. |
| Roderick MacKinnon Howard Hughes Medical Institute Laboratory of Molecular Neurobiology and Biophysics Rockefeller University 1230 York Avenue, New York, New York 10021 USA |
US citizen. Born 1956 (47 years). Grew up in Burlington outside Boston, USA. B.A. at Brandeis University, Boston, 1978. M.D. at Tufts Medical School in Boston, 1982. Since 1996, Professor of Molecular Neurobiology and Biophysics at The Rockefeller University in New York. |